
领域前沿·中国
肌肉干细胞对骨骼肌健康的多重调控
胡苹
(1广州国家实验室, 广州 510005; 2广州医科大学附属第五医院广东省生物靶向与诊治与康复工程技术研究中心, 广州 510005)
 | 胡苹, 广州国家实验室研究员、博士生导师, 入选中国科学院BR计划、上海市重大人才计划、广东省重大人才计划。主要研究领域为: 组织干细胞与成体组织再生、病变及其在再生医学中的临床应用, 代表性成果发表于Cell、Nat Commun、Cell Res、EMBO J、Dev Cell等学术期刊。在国际上首次进行父源肌肉干细胞治疗杜氏肌营养不良的临床试验, 证实其安全性与有效性, 探索杜氏肌营养不良、肌少症、特发性脊柱侧弯等疾病的再生医学治疗新路径。 |
肌肉干细胞对骨骼肌健康的多重调控
胡苹1,2*
(1广州医科大学附属第五医院广东省生物靶向与诊治与康复工程技术研究中心, 广州 510005; 2广州国家实验室, 广州 510005)
【摘要】肌肉干细胞是存在于成体骨骼肌中的一种组织干细胞, 能够特异性分化为肌管细胞,从而实现对骨骼肌损伤的修复, 是骨骼肌损伤修复再生的主要执行者。肌肉干细胞具有较高的异质性, 其命运转化受到微环境因子和细胞内源转录与表观遗传网络的精密调控。随着更多准确模拟患者表型的动物模型的建立, 在这些模型中的新研究结果表明肌肉干细胞病变是很多骨骼肌疾病发生和发展的重要原因, 同时肌肉干细胞功能退变也是青少年特发性脊柱侧弯等传统意义上的非骨骼肌疾病的发生与发展的重要原因。因此, 建立有功能的肌肉干细胞的体外长期扩增体系对于通过细胞治疗的方式治疗肌肉干细胞相关的骨骼肌和非骨骼肌疾病有重要意义。
【关键词】肌肉干细胞; 再生; 骨骼肌疾病
The Multiple Roles of Muscle Stem Cells in Skeletal Muscle Health
HU Ping1,2*
(1Key Laboratory of Biological Targeting Diagnosis,Therapy and Rehabilitation of Guangdong Higher Education Institutes, the Fifth Affiliated Hospital of Guangzhou Medical University, Guangzhou 510005, China; 2Guangzhou Laboratory, Guangzhou 510005, China)
【Abstract】Muscle stem cells are a group of tissue stem cells present in skeletal muscles. Muscle stem cells can specifically differentiate to myotubes and support the regeneration of skeletal muscles. Muscle stem cells are the major executors of muscle regeneration. They are highly heterogeneous and regulated by a well knitted network of microenvironment and cell intrinsic molecular signaling. Not only many skeletal diseases but also nonmuscle diseases like adolescent idiopathic scoliosis turn out to be caused by the dysfunction of muscle stem cells. Therefore, muscle stem cell transplantation will improve and cure these stem cell related diseases. The establishment of the in vitro expansion system to expand functional muscle stem cells has great impacts on the development of cell therapies for these muscle stem cell related diseases.
【Keywords】muscle stem cells; regeneration; muscle diseases
1 肌肉干细胞与骨骼肌再生
骨骼肌是体内最大的组织器官, 不仅能够通过收缩产生力, 支持集体的主动运动、姿态维持及呼吸, 而且具有重要的代谢和免疫调节功能。骨骼肌具有较强的再生能力, 能够高效修复日常活动导致的骨骼肌磨损及拉伤、扭伤等急性骨骼肌损伤[1-2]。骨骼肌的再生主要是由肌肉干细胞(muscle stem cells, MuSCs)执行的。MuSCs是存在于成体骨骼肌中的组织干细胞, 具有自我更新能力, 能够特异性地分化为肌管细胞, 从而实现对骨骼肌损伤的修复[3]。
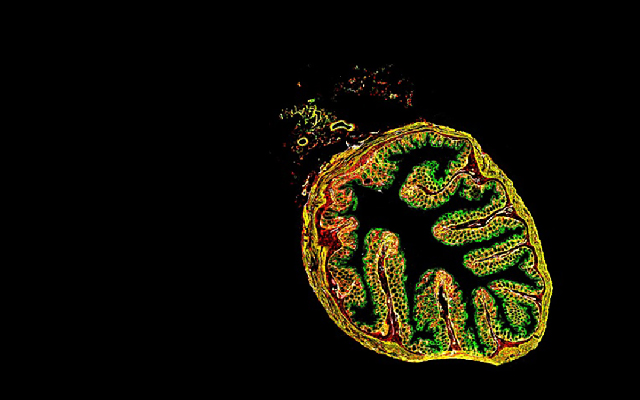
肌肉干细胞处在肌膜(membrane of myofibers)和基底膜(basal membrane)之间的干细胞巢(niche)中(图1)[4], 处于G0期的静息(quiescence)状态。在骨骼肌稳态下, 干细胞巢中的胞外基质(extracellular matrix, ECM)、多种分泌因子等共同构成稳定的微环境, 维持肌肉干细胞的静息[1,5]。在骨骼肌稳态时,MuSCs所处静息态的深浅程度并不相同, 有较强的异质性(heterogeneity)。一群表达Gli1的MuSCs处在更浅的静息状态, 在损伤发生时首先被动员[6]。损伤发生后, MuSCs被激活回归细胞周期, 进入细胞周期后, MuSCs经过短暂扩增, 分化为成肌细胞(myoblasts),成肌细胞经过扩增后, 一方面可以与受损肌纤维融合, 进行损伤修复; 另一方面, 也可以独立分化为肌管细胞(myotubes)后形成新的肌纤维, 完成损伤修复。同时, MuSCs也重新回到静息态, 为下一次的损伤修复做准备[1,5](图2)。这一过程贯穿整个生命周期, 对保持健康的骨骼肌至关重要。
绝大多数MuSCs表达paired box家族转录因子Pax7。Pax7被认为是MuSCs的最重要的标志基因。有少量MuSCs不表达Pax7, 而表达同属paired box家族的转录因子Pax3。在正常骨骼肌损伤修复中, 由于Pax3+ MuSCs细胞数量很小, 因此其对再生的贡献非常小。但是Pax3+ MuSCs具有较低的内源活性氧(reactive oxygen species, ROS)水平, 在辐射等恶劣环境下具有更强的增殖能力[7-8]。Pax3+ MuSCs可能是在进化中被选择出来在体内用于在恶劣环境下保持骨骼肌再生能力, 维护骨骼肌功能的干细胞。

图1 肌肉干细胞在骨骼肌中的位置(引自参考文献[4])
Fig.1 The localization of muscle stem cells (cited from reference [4])

图2 肌肉干细胞在再生中的命运转变
Fig.2 The conversion of muscle stem cell fate during regeneration
2 肌肉干细胞的活性调控
在骨骼肌稳态和损伤修复过程中, MuSCs从静息到激活, 从增殖到分化, 每一步命运改变都受到严格的调控。参与调控MuSCs命运转变的既包括多种微环境细胞和微环境因子, 也包括干细胞的多种内源信号通路。
损伤发生时, 不仅损伤部位的物理和化学微环境会发生显著的改变, 而且还会有来自损伤部位的因子通过血管被运送到全身, 产生远程效应。例如,损伤小鼠后腿一侧骨骼肌之后, 损伤部位会产生肝生长因子激活因子(hepatocyte growth factor activator,HGFA), 远程调控对侧腿骨骼肌中的MuSCs, 激活其中的mTOR信号通路使其进入GAlert状态, 更快地响应损伤信号离开静息态激活[9]。对于Gli1+MuSC亚群的进一步研究表明, 感受对侧损伤信号进行响应的主要是Gli1+ MuSCs, GAlert主要来源于Gli1+MuSCs[6]。
损伤发生后, MuSCs保持静息的微环境被破坏。损伤既会破坏肌膜和基底膜, 从而改变MuSCs周围的ECM种类和结构, 还会由于血管的破裂导致大量炎症细胞的浸润。除了细胞和ECM微环境的变化外,损伤还会导致大量细胞死亡, 从而导致多种细胞内容物的释放, 同时, 各种微环境细胞也会分泌多种因子, 造成MuSCs周围的化学微环境发生重大改变, 从而与细胞内源信号网络共同改变MuSCs的命运。
在急性损伤发生的早期, 肌纤维受损后会发生大量死亡。这些受损肌纤维发生程序性坏死(necroptosis)。发生程序性坏死的肌纤维激活TNC(Tenasin C)的表达, 随着程序性坏死细胞膜的破裂,TNC随其他细胞内容物一起被释放到损伤部位, 进入MuSCs的微环境。TNC的N-端存在类EGF结构域(EGF like domain), 这一结构域可以模拟EGF与MuSCs的EGF受体(EGF receptor, EGFR)结合, 进而激活EGFR信号通路, 促进MuSCs的增殖[10]。
受损肌纤维的存在还会激活免疫系统, 造成大量炎症细胞的浸润, 从而诱发急性炎症反应。这类在再生过程中发生的无菌炎症与感染诱发的炎症有很大的区别, 无菌炎症在骨骼肌再生中发挥重要的调控作用[1-2,10-11]。巨噬细胞在骨骼肌再生过程中分泌多种因子调控MuSCs从激活、增殖到分化的多种命运转换[1-2,11-12]。T细胞在损伤发生后被大量招募至损伤部位并被激活。由T细胞介导的炎症是骨骼肌再生所必需的, 缺乏T细胞的免疫缺陷型小鼠表现出明显的骨骼肌再生障碍, 移植T细胞可以纠正免疫缺陷小鼠的骨骼肌再生障碍, 说明T细胞在骨骼肌再生中扮演不可或缺的角色。进一步的研究发现,T细胞分泌的四种炎症因子(IL-1α、IL-13、TNFα、IFNγ)组合能够保持MuSCs的干性, 同时促进MuSCs的增殖, 实现具有干性的MuSCs的扩增[13]。大部分组织干细胞都难以在体外进行功能性扩增, 在体外培养后的组织干细胞或者无法有效扩增, 或者在扩增过程中丢失干性, 无法在体内支持再生。因此阐明体内组织干细胞的增殖原理对于建立组织干细胞在体外的高效功能性扩增体系具有重要的指导意义。
基于对MuSCs体内免疫微环境的研究, 在体外MuSCs扩增体系中加入IL-1α、IL-13、TNFα、IFNγ四种炎症因子组合后, MuSCs在体外培养体系中的增殖能力显著增强。MuSCs在体外培养超过24小时后会发生干性丢失, 移植入体内后丧失在体内再生为肌纤维的能力。在加入四种炎症因子后, MuSCs可以在体外长期传代至p40, 每一代MuSCs均具有MuSCs的分子特征, 在移植入体内后均能够高效支持对急性损伤的修复。在体外长期扩增的MuSCs移植入体内之后, 能够进入体内的干细胞库, 进入静息状态。在体外长期扩增后的MuSCs在移植后不仅能够支持对移植时损伤的修复, 而且在发生后续骨骼肌损伤时, 进入体内干细胞库的MuSCs可以被再次激活, 支持对后续损伤的修复。加入四种炎症因子后培养的MuSCs在体内具有干细胞的全部性质, 说明通过模拟体内MuSCs的免疫微环境, 在体外可以高效扩增保持完整干性的有功能的MuSCs, 为骨骼肌损伤和骨骼肌疾病的细胞治疗提供足量的优秀种子细胞[13]。
在受到微环境的调控的同时, MuSCs的增殖、分化等命运转换还受到复杂的表观遗传和转录调控。bHLH(basisc helix-loop-helix)家族转录因子MyoD长期以来被认为是肌向分化的核心转录因子[3,5], 能够进一步激活分化关键转录因子Myogenin的表达。Myogenin同样属于bHLH转录因子家族, 能够激活终末分化关键基因Myh1、MCK等的表达[1]。FoxO3能够与Pax7相互作用, 促进MuSCs中MyoD的表达,从而起始MuSCs的分化[14]。对全基因组转录因子的分析表明, FoxO3与MyoD共同决定MuSCs的超级增强子[15], 在MuSCs命运转换中发挥多面功能。FoxO3既能够在静息态MuSCs中促进静息, 抑制激活[16]; 又能够在激活的MuSCs中促进MyoD表达, 促进MuSCs分化[14]。转录辅因子Vgll4在MuSCs增殖时期作为Yap信号通路的抑制因子, 通过与YAP竞争性结合TEAD4转录因子, 抑制YAP靶基因CTGF等的转录, 防止MuSCs过度增殖。在分化起始时, Vgll4转变为转录激活辅因子, 通过与TEAD4和MyoD形成三元复合物, 激活Myogenin的转录[17]。
Myogenin基因的转录还受到DNA甲基化的调控。DNA双加氧酶Tet2可以通过氧化Myogenin增强子中MyoD结合位点附近的甲基化CpG岛, 进而降低MyoD结合位点附近的DNA甲基化水平, 从而稳定MyoD的结合。通过这一机制, Myogenin基因可以实现稳定的转录激活[18]。
MuSCs的命运转变还受到多种非编码RNA的调控。6种微小RNA(microRNAs, miRs) miR-1、miR-133a/b、miR-206、miR-208b、miR-499在骨骼肌中特异性高表达, 被称为myoMiRs, 调控骨骼肌的胚胎发育与成体骨骼肌中MuSCs的增殖与分化[19]。其他miRs在MuSCs中也发挥重要的调控作用[1]。这些miRs的表达水平受到宿主基因的转录及miRs的加工成熟等多方面的调控。例如, RNA结合蛋白Msi2与HuR共同抑制miR7a-1的成熟, miR7a-1通过结合节律基因Cry2 3′UTR序列抑制Cry2的翻译。Cry2能够促进MuSCs的分化, 因而Msi2和HuR这一对RNA结合蛋白通过双重抑制机制促进MuSCs的分化[20]。
除了miRs外, 长非编码RNA也对MuSCs的命运转变发挥重要的调控作用。长非编码RNA Lnc17000113A16RIK通过促进肌源转录因子MEF2D的翻译进而促进MuSCs的翻译[21]。长非编码RNAMyolncR4能够编码微肽LEMP, 促进MuSCs的分化[22]。
MuSCs的内源性调控机制涉及细胞活动的方方面面, 还需要很多深入研究, 阐明MuSCs从静息到激活、从激活到增殖、从增殖到分化的多重命运转化的精密调控机制。
3 肌肉干细胞与骨骼肌衰老和疾病
MuSCs不仅在骨骼肌损伤修复再生中作为再生的基本执行细胞发挥重要的作用, 而且也是骨骼肌衰老与疾病发生、发展的关键细胞群体。骨骼肌的衰老包括骨骼肌质量的降低、肌力等功能的衰减、再生能力减弱等。增龄导致的骨骼肌功能病态功能蜕变称为肌少症(sarcopenia)。目前肌少症的成因尚不清楚, 缺乏准确、客观、简便的早期诊断方法和有效的治疗方法。衰老的骨骼肌高表达分泌因子Dkk3, Dkk3能够抑制骨骼肌细胞中的Akt信号通路, 促进FoxO3和ꞵ-Catenin入核, 新入核的FoxO3与ꞵ-Catenin相互作用, 共同激活骨骼肌特异性E3泛素连接酶Atrogin 1和MurF 1的表达, 从而加速骨骼肌的萎缩。由于Dkk3是分泌因子, 因此一块骨骼肌的衰老可以诱导多块肌肉的衰老, 在全身“传染”[23]。除了肌肉萎缩之外, 由于MuSCs的增殖和分化能力减弱, 衰老骨骼肌的再生能力显著降低。衰老MuSCs的功能减弱受到多种因素的调控, 例如由于细胞自噬水平降低造成ROS的累积[24]。来源于衰老骨骼肌的MuSCs中的Msi2表达水平降低, 从而使得Msi2对miR-7a-1成熟的抑制能力降低, 提高成熟miR-7a-1在MuSCs中的表达水平, 抑制MuSCs的分化, 造成衰老骨骼肌的再生能力减弱[20]。针对这些衰老因素进行靶向干预能够延缓和逆转骨骼肌的衰老。例如, 通过RNAi敲低Dkk3的表达能够在衰老小鼠中有效地改善骨骼肌功能, 提高骨骼肌质量, 逆转骨骼肌衰老, 是潜在的肌少症治疗方法[23]。
很多骨骼肌疾病也与MuSCs功能障碍密切相关。强直性肌营养不良(myotonic dystrophy, DM1)是一种多系统遗传疾病, 影响骨骼肌、平滑肌和心肌, 患者肌肉强直、无力、吞咽困难、心律失常、学习困难、发生白内障等, 目前没有有效的治疗药物。这一疾病是常染色体隐性遗传疾病, 由DMPK(myotonic dystrophy protein kinase)基因3′UTR区域的CTG重复序列异常扩增造成。在小鼠DMPK基因的3′UTR区域添加80个CTG重复单元模拟患者的基因突变获得的DM1疾病模型仅能够模拟患者的少数轻微症状, 无法模拟临床的复杂多系统病变。应用单倍体细胞“人造精子”技术构建DMPK及其邻近基因SIX5、DMWD、MBNL1的半敲除杂合小鼠, 能够模拟DM1临床患者的全部多系统病变, 揭示DMPK 3′UTR区域CTG重复序列的异常扩增导致染色质结构异常, 干扰DMPK基因及其邻近基因的表达。这一染色质结构异常导致的多基因表达异常使DM1多系统病变发生。应用这一新型DM1疾病模型小鼠, 发现DM1 MuSCs分化异常, 首次模拟出DM1患者中的MuSCs功能异常, 为DM1治疗药物开发提供了新的靶细胞[25]。
杜氏肌营养不良(Duchenne muscular dystrophy,DMD)是由Dystrophin单基因突变导致的伴X隐性遗传疾病, 在男孩中的发病率为1/3 500。DMD主要表现为骨骼肌从远端向躯干中心的渐进性萎缩,在疾病晚期因呼吸系统并发症或心力衰竭而死亡。DMD长期以来被认为是由于终末分化的骨骼肌损毁而导致的疾病。通过模拟Dystrophin基因缺失制备了小鼠、狗、猪等动物模型, 这些模型在疾病机制研究、新药评价等工作中广泛应用[26-28]。但是,这些动物模型中DMD的进程与人类患者中的情况相去甚远, 使得对疾病进展的机制及药物评价的准确性不高。非人灵长类动物与人类在基因组序列上最为接近, 在生理、解剖、发育方面也高度相似, 能够更好地模拟人类疾病。通过CRSPR/Cas9介导的基因编辑技术, 科学家制备了DMD非人灵长类猕猴模型[29]。该模型在疾病发生的非常早期即可模拟DMD患者的下肢无力、表征肌肉损毁的血清学指标上调、步态改变等行为学特征, 以及肌肉严重纤维化、肌肉损毁等组织学表型。在DMD早期的非人灵长类模型中, 通过骨骼肌单细胞测序分析发现骨骼肌中有大量免疫细胞浸润, 与临床观察中看到的患者骨骼肌中免疫细胞增多, 发生炎症等症状相吻合[29]。对于骨骼肌中的FAPs(fibro-adipo progenitors)的研究表明, 与小鼠中的情况不同, 虽然骨骼肌发生了严重的纤维化, 但是FAPs细胞并没有大量增殖[29]。DMD非人灵长类模型中, FAPs细胞亚群之间的谱系关系紊乱, 同时所有亚群中纤维化相关基因的表达都显著上调, 提示DMD发病早期骨骼肌的纤维化可能是由于FAPs细胞中普遍的纤维化基因上调导致的。TGFꞵ信号通路是目前激活纤维化基因表达的主要信号通路。但是, DMD早期骨骼肌中的纤维化却不受TGFꞵ信号通路的调控。长期以来, DMD的发病机制一直被认为是肌纤维的持续损毁。持续的肌纤维损毁会激活MuSCs, 发生持续的骨骼肌损伤修复, 导致MuSCs耗竭。而在非人灵长类DMD模型中的研究则表明, MuSCs在疾病的早期即已经发生显著的细胞内源性改变, 细胞凋亡的比例升高, 表现出明显的细胞内源性分化障碍[29]。因此, DMD不仅是发生在分化肌纤维中的肌肉损毁的疾病, 同时也是一种干细胞疾病[29]。治疗DMD不仅要修正肌纤维中的问题, 而且要修正MuSCs的功能障碍, 为DMD的治疗提供了新的路径。
MuSCs不仅在传统的骨骼肌疾病中发挥重要的功能, 而且还在一些非骨骼肌疾病的进展中发挥作用。青少年特发性脊柱侧弯(adolescent idiopathic scoliosis, ALS)是多发于青春期女性中的疾病, 其发病原因不清楚。我们的研究表明, ALS中凹侧椎旁肌中的MuSCs中ESR1的表达水平显著下调, 进而导致ESR1信号通路下调。而凸侧椎旁肌中的MuSCs中ESR1的表达水平保持正常。ESR1信号通路水平下调导致MuSCs分化障碍。两侧椎旁肌中MuSCs功能的不对称导致凹侧椎旁肌的肌纤维直径变小, 力量减弱, 无法为脊柱提供平衡的支撑力量, 造成脊柱侧弯。在小鼠模型中通过抑制一侧椎旁肌中的ESR信号通路可以加速AIS的进展。在AIS小鼠模型的凹侧椎旁肌中通过注射FDA批准的药品雷洛昔芬能够重新激活ESR信号通路, 恢复MuSCs的分化能力,从而显著减缓AIS的进展[30]。这一工作揭示了肌肉力学支撑对于保持正常骨骼功能的重要作用, 为骨骼疾病的治疗提供了新的思路。
最近的研究表明, 除了分化为骨骼肌外, 人MuSCs还具有肌腱分化潜能。在损伤的肌腱中移植MuSCs, 能够显著促进肌腱的修复, 改善运动功能[31]。肌腱损伤难以修复, 肌腱中虽然有肌腱干细胞/前体细胞存在, 但是由于其数量非常少, 在体外几乎无法扩增, 因而很难支撑再生医学应用。应用我们建立的功能MuSCs长期扩增体系, 能够获得足量优质的MuSCs, 以用于肌腱损伤修复, 为肌腱损伤的再生医学治疗提供了新的种子细胞。
MuSCs在多种疾病中的功能, 及针对MuSCs开发治疗方法尚需进一步深入研究。借助功能MuSCs体外长期阔体系通过干细胞移植进行细胞治疗等新的治疗方法为很多骨骼肌和非骨骼肌疾病的治疗带来了新的希望。
4 展望与思考
MuSCs作为一种组织干细胞通过增殖分化直接支持骨骼肌的再生, 在骨骼肌的再生和稳态维持中发挥决定性的作用。MuSCs的研究和临床应用都需要面对MuSCs的异质性问题。MuSCs经历静息–激活–增殖–分化, 及静息态时的自我更新等多种细胞状态的转变, 单细胞测序的结果表明, 在再生过程中存在着很多处于上述状态之间的中间态细胞[32-33],使得细胞状态的异质性更为复杂。同时, 在MuSCs中还存在细胞内源性异质性, 例如Gli1+ MuSCs是一群处在更浅的静息态的MuSCs, 但是大多数静息态MuSCs在激活过程中并不经过Gli1表达的过程[6], 即Gli1+ MuSCs并不是静息态细胞激活过程中产生的中间态, 而是一个有独特基因表达模式的MuSCs亚群。在体内, 细胞状态的多样性和源于细胞内源性差异的亚群多样性组合在一起, 使得MuSCs具有很高的异质性。在疾病情况下, 异质性进一步加强[29]。进一步解析MuSCs的异质性及其驱动机制, 一方面可以加深对MuSCs细胞命运转变的路径选择的理解; 另一方面, 对于MuSCs异质性的研究有助于鉴定具有更高干性和更强增殖能力的MuSCs, 为MuSCs的临床应用提供更优的种子细胞。
传统上MuSCs主要在骨骼肌损伤修复中发挥作用, 骨骼肌的衰老与多数疾病更多情况下被认为是终末分化的肌纤维的代谢、蛋白稳态被破坏, 发生功能退变而发生的。通过制备能够更准确地模拟患者表型的多种动物模型, 使得获取充足的骨骼肌样品进行深入研究成为可能。这些研究表明,MuSCs的病变在多种骨骼肌疾病的发病及进展中都发挥重要的作用。因此, 在治疗骨骼肌病变的时候,对MuSCs的功能校正也是治疗的重要考虑。此外,MuSCs的功能退变在一些非骨骼肌疾病的发生、发展中也发挥重要的作用。因此, 校正和增强MuSCs的功能不仅有望治疗骨骼肌疾病, 也可以用于非骨骼肌疾病的治疗。
由于功能MuSCs体外长期扩增体系的建立,我们能够获得足量的优质MuSCs, 因此通过健康MuSCs移植或在MuSCs中进行基因编辑校正基因缺陷从而实现对骨骼肌疾病的治疗都成为可能。建立高效移植系统、追踪移植细胞在体内的功能及转归,进一步提高肌肉干细胞治疗的安全性和有效性都是骨骼肌疾病细胞治疗的重要研究方向。应用MuSCs进行疾病治疗有望治愈很多目前缺乏有效治疗方法的疾病。
参考文献 (References)
[1] FU X, ZHUANG C L, HU P. Regulation of muscle stem cell fate [J]. Cell Regen, 2022, 11(1): 40.
[2] YANG W, HU P. Skeletal muscle regeneration is modulated by inflammation [J]. J Orthop Translat, 2018, 13: 25-32.
[3] FU X, HE Q, TAO Y. et al. Recent advances in tissue stem cells [J]. Sci China Life Sci, 2021, 64(12): 1998-2029.
[4] 刘晟, 胡苹. 类器官及其应用第7章3D骨骼肌类器官[M]. 上海: 上海科学技术出版社, 2023.
[5] FU X, WANG H, HU P. Stem cell activation in skeletal muscle regeneration [J]. Cell Mol Life Sci, 2015, 72(9): 1663-77.
[6] PENG J, HAN L, LIU B, et al. Gli1 marks a sentinel muscle stem cell population for muscle regeneration [J]. Nat Commun, 2023, 14(1): 6993.
[7] DER VARTANIAN A, QUÉTIN M, MICHINEAU S. et al. PAX3 confers functional heterogeneity in skeletal muscle stem cell responses to environmental stress [J]. Cell Stem Cell, 2019, 24(6): 958-73.
[8] SCARAMOZZA A, PARK D, KOLLU S, et al. Lineage tracing reveals a subset of reserve muscle stem cells capable of clonal expansion under stress [J]. Cell Stem Cell, 2019, 24(6): 944-57.
[9] RODGERS J T, KING K Y, BRETT J O, et al. mTORC1 controls the adaptive transition of quiescent stem cells from G0 to G(Alert) [J]. Nature, 2014, 510(7505): 393-6.
[10] ZHOU S, ZHANG W, CAI G, et al. Myofiber necroptosis promotes muscle stem cell proliferation via releasing Tenascin-C during regeneration [J]. Cell Res, 2020, 30(12): 1063-77.
[11] YANG W, HU P. Hierarchical signaling transduction of the immune and muscle cell crosstalk in muscle regeneration [J]. Cell Immunol, 2018, 326: 2-7.
[12] PANCI G, CHAZAUD B. Inflammation during post-injury skeletal muscle regeneration [J]. Semin Cell Dev Biol, 2021, 119: 32-8.
[13] FU X, XIAO J, WEI Y, et al. Combination of inflammationrelated cytokines promotes long-term muscle stem cell expansion [J]. Cell Res, 2015, 25(6): 655-73.
[14] HU P, GELES K G, PAIK J H, et al. Codependent activators direct myoblast-specific MyoD transcription [J]. Dev Cell, 2008,15(4): 534-46.
[15] PENG X L, SO K K, HE L, et al. MyoD- and FoxO3-mediated hotspot interaction orchestrates super-enhancer activity during myogenic differentiation [J]. Nucleic Acids Res, 2017, 45(15): 8785-805.
[16] GOPINATH S D, WEBB A E, BRUNET A, et al. FOXO3 promotes quiescence in adult muscle stem cells during the process of self-renewal [J]. Stem Cell Reports, 2014, 2(4): 414-26.
[17] FENG X, WANG Z, WANG F, et al. Dual function of VGLL4 in muscle regeneration [J]. EMBO J, 2019, 38(17): e101051.
[18] WANG H, HUANG Y, YU M, et al. Muscle regeneration controlled by a designated DNA dioxygenase [J]. Cell Death Dis, 2021, 12(6): 535.
[19] HORAK M, NOVAK J, BIENERTOVA-VASKU J. Muscle-specific microRNAs in skeletal muscle development [J]. Dev Biol, 2016, 410(1): 1-13.
[20] YANG W, YANG L, WANG J, et al. Msi2-mediated MiR7a-1 processing repression promotes myogenesis [J]. J Cachexia Sarcopenia Muscle, 2022, 13(1): 728-42.
[21] FU X, LI S, JIA M, et al. Myogenesis controlled by a long noncoding RNA 1700113A16RIK and post-transcriptional regulation [J]. Cell Regen, 2022, 11(1): 13.
[22] WANG L, FAN J, HAN L, et al. The micropeptide LEMP plays an evolutionarily conserved role in myogenesis [J]. Cell Death Dis, 2020, 11(5): 357.
[23] YIN J, YANG L, XIE Y, et al. Dkk3 dependent transcriptional regulation controls age related skeletal muscle atrophy [J]. Nat Commun, 2018, 9(1): 1752.
[24] GARCÍA-PRAT L, MARTÍNEZ-VICENTE M, PERDIGUERO E, et al. Autophagy maintains stemness by preventing senescence [J]. Nature, 2016, 529(7584): 37-42.
[25] YIN Q, WANG H, LI N, et al. Dosage effect of multiple genes accounts for multisystem disorder of myotonic dystrophy type 1 [J]. Cell Res, 2020, 30(2): 133-45.
[26] BULFIELD G, SILLER W G, WIGHT P A, et al. X chromosome- linked muscular dystrophy (mdx) in the mouse [J]. Proc Natl Acad Sci USA, 1984, 81(4): 1189-92.
[27] COOPER B J, WINAND N J, STEDMAN H, et al. The homologue of the Duchenne locus is defective in X-linked muscular dystrophy of dogs [J]. Nature, 1988, 334(6178): 154-6.
[28] KLYMIUK N, BLUTKE A, GRAF A, et al. Dystrophin-deficient pigs provide new insights into the hierarchy of physiological derangements of dystrophic muscle [J]. Hum Mol Genet, 2013, 22(21): 4368-82.
[29] REN S, FU X, GUO W, et al. Profound cellular defects attribute to muscular pathogenesis in the rhesus monkey model of Duchenne muscular dystrophy [J]. Cell, 2024, 187(23): 6669-86,e16.
[30] SHAO X, FU X, YANG J, et al. The asymmetrical ESR1 signaling in muscle progenitor cells determines the progression of adolescent idiopathic scoliosis [J]. Cell Discov, 2023, 9(1): 44.
[31] SHAO X X, LIN X Z, ZHOU H, et al. Human myogenic progenitor cells display tenogenic differentiation potential and facilitate tendon regeneration [J]. eLife, 2024, https://doi.org/10.7554/eLife.98636.1.
[32] OPRESCU S N, YUE F, QIU J, et al. Temporal dynamics and heterogeneity of cell populations during skeletal muscle regeneration [J]. 2020, 23(4): 100993.
[33] DELL’ORSO S, JUAN A H, KO K D, et al. Single cell analysis of adult mouse skeletal muscle stem cells in homeostatic and regenerative conditions [J]. Development, 2019, 146(12): dev174177.

 中文版
中文版 英文版
英文版